

Translate this page into:
Hepatocyte nuclear factor-1β: A regulator of kidney development and cystogenesis
This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-Share Alike 3.0 Unported, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This article was originally published by Medknow Publications & Media Pvt Ltd and was migrated to Scientific Scholar after the change of Publisher.
Abstract
The understanding of the genomics of the renal tissue has gathered a considerable interest and is making rapid progress. The molecular mechanisms as well as the precise function of the associated molecular components toward renal pathophysiology have recently been realized. For the cystic kidney disease, the regulation of gene expression affecting epithelial cells proliferation, apoptosis as well as process of differentiation/de-differentiation represent key molecular targets. For the cystic disorders, molecular targets have been identified, which besides lending heterogeneity to cysts may also provide tools to unravel their functional importance to understand the renal tissue homeostasis. This review focuses on providing comprehensive information about the transcriptional regulatory role of hepatocyte nuclear factor-1β, a homeoprotein, as well as its interacting partners in renal tissue development and pathophysiology.
Keywords
Kidney development
hepatocyte nuclear factor-1β
renal cystogenesis
transcription factor

Introduction
Kidneys play a crucial role in maintaining fluid homeostasis and excreting waste products. Changes that affect kidney functions are reflected by altered renal filtration system, often leading to end-stage renal disease (ESRD). Today, millions of individuals globally suffer from ESRD and need renal transplant therapy.[1]
Kidney has been used as a classical model to study tissue morphogenesis. Genetic disorders of kidney reflecting abnormal development have been studied in conjunction with gene expression and regulation. In this regard, understanding the role of both the trans-acting factors and cis-binding element is important. The transcription factors maintain the fine regulatory control both for the tissue development and pathophysiology.[2] In recent years, these have been targeted as potential therapeutic targets for certain disease.[3]
Cell culture experiments and animal models have helped in identification of novel transcription factors for kidney development in maintaining homeostasis.[45] Role of transcription factors varies in different tissues depending on their development, whether early or later at stage. With respect to eukaryotes, transcription involves a complete array of different and specific interactions among nuclear proteins and DNA elements located in the promoter or enhancer region of genes.[6] Transcription factors being limited to specific types of lineage or to a whole tissue activate various genes sequentially during kidney development, thereby sustaining the integrity of a normal kidney.[7] Many transcription factors may be mutated or have abnormal expression causing congenital abnormalities of the kidneys such as renal agenesis, cysts, hypoplasia, dysplasia and ureteric malformations [Table 1].

Hepatocyte nuclear factor-1 (HNF-1), a nuclear protein, was first identified in hepatocytes as a regulator of gene transcription of β chain of fibrinogen.[22] It represents a family of transcription factors consisting of HNF-1α and HNF-1β. Both these factors have been characterized as mutual nuclear proteins interacting with a single crucial element of the albumin promoter. HNF-1α was found only in liver or differentiated hepatoma cells, while HNF-1β was present in dedifferentiated hepatoma cells as well as in somatic cell hybrids that showed extinction of liver-specific proteins, including albumin.[23] However, their role during kidney development and in cystogenesis through regulation of cystic genes have revealed the importance of these proteins in renal tissue homeostasis.
Hepatocyte Nuclear Factor-1: Structure and Domains
HNF-1 family members, HNF-1α and HNF-1β, share conserved N-terminal dimerization domain structure, a Pit-1/Oct-1/Unc-86 (POU) domain and a homeodomain for DNA binding and a less-conserved C-terminal dimerization domain allowing binding of homo- or hetero-dimerization of the HNF-1 proteins.[24]
HNF-1α is expressed as a single 631 amino acid polypeptide located at chromosome 12, and HNF-1β is expressed as two polypeptides having 557 amino acids placed on chromosome 17.[25] First amino acids of the polypeptide chain of the proteins promote dimerization of the two proteins during transcription activation. POU domain (55–60 amino acids), which cultivates DNA binding, is located adjacent to the dimerization domain [Figure 1]. Both of these interacting partners either work independently of each other as homodimers or work in specific ratios as heterodimers in regulating their concerned genes.[26] The genes for HNF-1s have been highly conserved throughout vertebrate evolution.[27] Extensive conservation is seen in the dimerization domain and DNA binding domain, but both the proteins differ in their thermostability and their molecular weight, even though they display same sequence specificities often recognizing the same consensus palindromic sequence 5'-GTTAATNATTAAC-3'[282930] when forming heterodimers. Moreover, there are reports suggesting against the strict requirement for the palindromic sequence for binding of HNF-1α to the promoter region of genes from different organisms.[31]
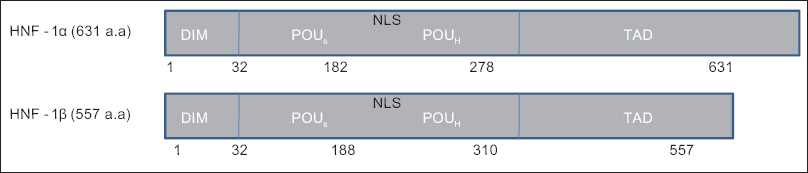
- Hepatocyte nuclear factor-1 (HNF-1) structure and domains. HNF-1α is expressed as a single polypeptide of 631 amino acids and HNF-1 β is expressed as two polypeptides of 557 amino acids. HNF-1: Hepatocyte nuclear factor-1; DIM: Dimerization domain; POUs: Pit-1, Onc and Unc specific domain; POUH: Pit-1, Onc and Unc-homeodomain; TAD: Transactivation domain; NLS: Nuclear localization signal
Hepatocyte Nuclear Factor-1β Expression During Early Development
HNF-1β has attracted attention for its role in early development and nephrogenesis as HNF-1α remains confined to hepatocyte development. It has been demonstrated that between the two proteins, the expression of HNF-1β precedes that of HNF-1α gene during embryogenesis, appearing at embryonic day 10.5 (E10.5).[323334] Although the endodermally derived structures, the foregut and the hindgut, lack messenger RNA (mRNA) expression, the midgut endodermally derived structures as well as the mesoderm-derived structures, such as the kidneys, express HNF-1β mRNAs. Inductive influence of HNF-1β along with its partners at certain stages may be required for organogenesis as they control the lateral plate mesoderm on the midgut endoderm.[35] Nullzygous mutant embryos have normal placement of parietal endoderm like cells on abnormal basal membrane, but show no activation of its other co-activators, HNF-4α, HNF-3α and HNF-1α, and die in utero. Many of the transient rodent models also showed earliest detection of HNF-1β mRNAs in the mouse primitive endoderm 4.5 days post-coitus followed by its expression in the yolk sac and developing gut.[3136] This reflects the action of HNF-1β in the primitive endoderm required for specification of visceral endoderm. Transgenic mice deficient in HNF-1β develop normally till blastocyst stage and die soon thereafter. Its inactivation in the mouse shows lethality in the embryo approximately at day 7.5 due to visceral endoderm differentiation and abnormal extra-embryonic region.[37] Both endodermal and mesodermal structures express HNF-1β pattern that reflects its involvement in embryonic segmental pattern. Many of the mice models devoid of HNF-1β, having restored its early expression using tetraploid embryo complementation, show normal pancreas morphogenesis and liver specification.[38]
Hepatocyte Nuclear Factor-1β: A Kidney Positive Transcription Factor
In vertebrates the pronephros, the mesonephros and the metanephros are formed progressively during development. Xenopus laevis is the most attractive model to analyze the molecular and cellular events in the pronephros formation. The genes for HNF-1s are highly conserved during the vertebrate evolution and show high-sequence conservation among species and also similarity to the frog, Xenopus.[3940] Expressed as an early marker of kidney development, HNF-1β is deeply involved in morphogenetic events. Overexpression of frameshift HNF-1β mutant P28L32fsdelCCTCT showed associated nephron agenesis in Xenopus larvae.[41] Likewise introduction of mutant form of HNF-1β into the developing frog embryo lead to partial or even complete agenesis of the pronephric tubules and duct. Pre-eminent position of HNF-1β in the regulatory network of differentiation during early embryogenesis reveals the importance of HNF-1β in developmentally regulated processes consistent with its early expression in primitive endoderm.
Kidney, an HNF-1β-positive tissue, expresses it early during development leading to activation of numerous genes involved in embryogenesis. Although HNF-1α is also expressed during the development of kidney, its role remains confined to genes transcribing glucose-6-phosphate transporter, and having significantly reduced mRNA levels.[42] The function and characterization of HNF-1β was first highlighted when human mutations were examined causing maturity-onset diabetes of the young (MODY).[4344] Therefore, its transcriptional role required for precise metabolic control was emphasized by analyzing insulin release by β-cells and hence, suggesting its wider role in influencing MODY5 due to added mutations. Although first identified as a "liver enriched" transcription factor, its role in kidney became apparent upon observation of renal abnormalities seen due to mutations and deficiency of HNF-1β in humans as well as experimental and transgenic animal models while studying MODY5. Its expression during kidney organogenesis reflects its importance during interaction between epithelium of the ureter and condensed mesenchymal cells.[45] HNF-1β is expressed in all segments of nephrons and renal collecting ducts[3746] and hence, crucial for kidney organogenesis. Moreover, identification of HNF-1β gene products to the primary cilium provided interesting observations as the deregulation of the primary ciliary processes are important for cystogenesis. Thus, many of the HNF-1β-targeted proteins, which also co-localize to primary cilium form important group of proteins for understanding cyst development and expansion. Although a predominant role of HNF-1β has been highlighted in recent years controlling kidney organogenesis and associated disease afflictions, more clear insight into the transcription role of HNF-1β in regulation of cystic genes, polycystic kidney disease (PKD) 2, PKHD1, UMOD, and KIF show its direct binding to their promoters. Most of these genes are co-localized to primary cilium as well. Transgenic mice lacking endogenous expression of HNF-1β showed cyst formation and down-regulation of PKD2, PKHD1, IFT88 (Polaris), and UMOD.[474849] HNF-1β transcriptional binding sites have been identified in the promoter regions of kidney-specific cadherin (ksp-cdh), cdh 16 and Na-K-Cl co-transporter,[50] existing mostly upstream of transcriptional start sites. Many of the kidney-specific expression of transporters involve coordinated regulation of HNF-1β and DNA methylation. The organic anion transporters (OATs) OAT1/SLC22A6, OAT3/SLC22A8 and urate transporter expressed predominantly in the kidney and placenta are directly transcribed by HNF-1β.[515253] Any of the mutant form of HNF-1β regulating the promoter regions and/or the OAT relate to the abnormal development of the kidney, leading to kidney deformities. Collectrin is also an important target of HNF-1β-mediated regulation. This protein is also localized to the cilium and involved in maintenance of the primary cilium processes that are important to cystogenesis. Knockouts of collectrin in mice resulted in the phenotype of diabetes, hypertension and renal cysts.[5455] Since HNF-1β mutant mice are also observed to produce renal cysts, its association with regulating promoter region of collectrin has shown its role toward cystogenesis.[56] Taken together, these observations provide convincing evidence that HNF-1β is at the center stage with diverse roles in ciliary functions and plays an important role in PKD.
As a transcription factor, it plays an important role in the primary development of various organs, including kidneys as one of its major target. In this regard, it has been shown that any of the mutant forms of HNF-1β regulating the promoter regions and/or the organic anion transporter relate to the abnormal development of the kidney and leading to kidney deformities.
Kidney Abnormalities and Cystogenesis Involving Hepatocyte Nuclear Factor-1β
HNF-1β and its correlated mutations were first described in patients with MODY. Screening revealed other manifestations, including cystic kidneys, genital malformations and kidney dysfunctions among the MODY5 patients,[255758]. Some of the mutations in HNF-1β are characterized with nondiabetic renal disease, and some show renal cysts as the major diagnosis.[59] Cystic diseases commonly surface in the screening of patients with diabetes, and many of the unrelated patients also carry heterozygous mutations for HNF-1β; having this associated disorder was termed as renal cysts and diabetes (RCAD).[6061] Screening of patients possibly carrying HNF 1β mutations causing renal cyst or nephron agenesis, but no diabetes is reportedly a gain of function mutation among the patients.[62]
Patients of varying age nowadays are investigated for HNF-1β mutations in order to analyze its wide spectrum in disease affliction, and not only limited to the kidneys. Prenatal diagnosis of a patient showed to develop bilateral hypoplastic kidneys at 3 months of age carrying heterozygous mutation (S148L) of HNF-1β gene and a compatible MODY5 phenotype requiring insulin, suggestive of nonautoimmune diabetes.[63] A 17-week-old fetus with this mutation lacked normal nephronogenesis with renal parenchyma replaced by cysts and occasional cystic glomeruli, including primitive tubules consistent with cystic renal dysplasia.[59] Another novel mutation recently was identified in a 12-year-old Romanian boy having progressive nondiabetic renal dysfunction and bilateral renal cyst formation. With the progress of HNF-1β mutations appearing collectively in these patients, new mutations are being categorized. A new mutations located at exon 3 (C715G7C p239R), was associated with renal insufficiency.[64] A 14-year-old boy presented nonketotic hyperglycemia, elevated serum creatinine levels and deranged liver function with no microalbuminuria or proteinuria. He showed mild mental retardation with learning difficulties. Ultrasonography of the abdomen revealed multiple renal cysts of various sizes in both the kidneys. Multiplex ligation-dependent probe amplification for dosage analysis of HNF-1β gene detected a heterozygous whole gene deletion (p.Met1_Trp557del) consistent with the diagnosis of RCAD syndrome. This is the first case of the RCAD syndrome reported in an Indian patient.[65] Many of the patients identified with E101X or P159Tdel mutations are also diagnosed with hypoplastic glomerulocystic kidney disease, and early onset diabetes[59] along with small kidneys with abnormal formation of calyces and papillae. MODY patients carrying A263insGG or P328L329del mutations often develop cystic dysplasia. Mutant carriers having R137-K161del[25] have reduced number of glomeruli and hypertrophy. A study of 109 patients for novel UMOD mutation in exon 8 causing familial juvenile hyperuricemic nephropathy and medullary cystic disease revealed six probands having HNF-1β.[66] Many of the patients carry a complete~1.4 Mb deletion at the chromosome 17q12.[67] Having observed complete deletion analysis, the screened Japanese patients also showed renal hypodysplasia (RHD) and unilateral multicystic dysplastic kidney. Deletion of HNF-1β in this part of the gene carries segmental breakpoints and microdeletions. Studies on patients carrying gene deletion in HNF-1β showed that majority of patients are found to have renal cystic disorders, leading to abnormal formation of kidney architecture.
Clinical Manifestations
Patient identified with MODY carrying R177X mutation showed additional renal defect.[45] Many other patients who were identified with the same malformation carried distinct features of diabetic nephropathy leading to microvascular complications in kidney and progressive increase in microalbuminuria, macroalbuminuria and renal failure.[68] Patients having HNF-1β gene mutations develop diabetes at a mean age of 17–25.8 years. Others (12–62.5%) develop genital malformations, rheumatic heart disease, pancreas atrophy, hyperuricemia and abnormal liver function. Other genital malformations such as bicornuate uterus and vaginal aplasia are usually seen in women, whereas asthenospermia, bilateral epididymal cysts and atresia of vas deferens are seen in men. Women with HNF-1β 17q12 deletion[69] show congenital aplasia of the uterus and upper part of the vagina.[707172] Patients with mutation of HNF-1β suffer from hypomagnesemic disorders where magnesium deficiency remains to be one of the most common electrolyte disorders. Families with a history of an inherited form of hypomagnesemia also show HNF-1β mutations, confirming its role in magnesium homeostasis in the distal convoluted tubules (DCTs) that control magnesium channel or the transient receptor channel, melastatin member 6,[73] including renal magnesium wasting. ChIP-chip data revealed HNF-1β transcriptional binding sites in the FXYD2 gene, which encodes γ-subunit of the Na+/K+-ATPase. The γ-subunit is one of the key molecular players in the renal Mg2+ reabsorption in DCT. HNF-1β inconsistency has been identified to cause congenital anomalies of the kidneys and urinary tract affecting bilateral renal malformations. Many patients identified with H69fsdelAC, H324S325fsdelCA, Y352finsA and K156E mutations suffered from hypomagnesemia. These mutations prevent the binding of HNF-1β transcription activation of γα-subunit.[7475]
The mechanisms controlling proliferation, differentiation and cell morphology of renal epithelial cells involve complex networks of interactive partners and transcription factors themselves having cross-talks on multiple levels. Transcription factors not only serve to regulate specific genes and cell populations, but their expressions coordinate the genetic programs and signaling pathways. Deciphering their role as cis-regulatory elements, they serve to create population of different renal cell types for normal functional kidney. In recent years, many of the transcription factors have been identified, but their role still remains elusive. In order to assess the functional role of HNF-1β in kidney development and the pathogenesis of the diseased kidneys, screening for HNF-1β mutations is important.
In-depth analysis of its functional significance as a transcription factor in kidney needs further analysis, and the possibility of mutated/truncated HNF-1β in patients with renal abnormalities should not be ruled out.
Source of Support: The work was supported by grant from Department of Biotechnology, (DBT), New Delhi. India to SP
Conflict of Interest: None declared.
References
- Global perspective of kidney disease. In: Byham-Gray LD, Burrowes JD, Chertow GM, eds. Nutrition in Kidney Disease. New Jersey: Humana Press; 2014. p. :11-3.
- [Google Scholar]
- Transcription factors and mammalian development. In: Pedersen RA, ed. Current Topics in Developmental Biology. San Diego, CA: Elsevier; 1992. p. :351-83.
- [Google Scholar]
- Transcriptional regulators in kidney disease: Gatekeepers of renal homeostasis. Trends Genet. 2008;24:361-71.
- [Google Scholar]
- GLI3-dependent transcriptional repression of Gli1, Gli2 and kidney patterning genes disrupts renal morphogenesis. Development. 2006;133:569-78.
- [Google Scholar]
- Transcription factor SIX5 is mutated in patients with branchio-oto-renal syndrome. Am J Hum Genet. 2007;80:800-4.
- [Google Scholar]
- Dynamics and interplay of nuclear architecture, genome organization, and gene expression. Genes Dev. 2007;21:3027-43.
- [Google Scholar]
- Distinct molecular and morphogenetic properties of mutations in the human HNF1beta gene that lead to defective kidney development. J Am Soc Nephrol. 2003;14:2033-41.
- [Google Scholar]
- Molecular genetics and mechanism of autosomal dominant polycystic kidney disease. Mol Genet Metab. 2000;69:1-15.
- [Google Scholar]
- The polycystic kidney disease proteins, polycystin-1, polycystin-2, polaris, and cystin, are co-localized in renal cilia. J Am Soc Nephrol. 2002;13:2508-16.
- [Google Scholar]
- Mutations in GPC3, a glypican gene, cause the Simpson-Golabi-Behmel overgrowth syndrome. Nat Genet. 1996;12:241-7.
- [Google Scholar]
- Mutations in the human Jagged1 gene are responsible for Alagille syndrome. Nat Genet. 1997;16:235-42.
- [Google Scholar]
- Epidermal growth factor receptor expression is abnormal in murine polycystic kidney. Kidney Int. 1995;47:490-9.
- [Google Scholar]
- TGF alpha overexpression in transgenic mice induces liver neoplasia and abnormal development of the mammary gland and pancreas. Cell. 1990;61:1137-46.
- [Google Scholar]
- Pax-2 controls multiple steps of urogenital development. Development. 1995;121:4057-65.
- [Google Scholar]
- Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature. 1994;372:679-83.
- [Google Scholar]
- The basic-helix-loop-helix protein pod1 is critically important for kidney and lung organogenesis. Development. 1999;126:5771-83.
- [Google Scholar]
- Fraser syndrome and mouse blebbed phenotype caused by mutations in FRAS1/Fras1 encoding a putative extracellular matrix protein. Nat Genet. 2003;34:203-8.
- [Google Scholar]
- PEX1 mutations in complementation group 1 of Zellweger spectrum patients correlate with severity of disease. Pediatr Res. 2002;51:706-14.
- [Google Scholar]
- Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell. 1994;79:1111-20.
- [Google Scholar]
- Purified hepatocyte nuclear factor 1 interacts with a family of hepatocyte-specific promoters. Proc Natl Acad Sci U S A. 1988;85:7937-41.
- [Google Scholar]
- A liver-specific factor essential for albumin transcription differs between differentiated and dedifferentiated rat hepatoma cells. Genes Dev. 1988;2:957-74.
- [Google Scholar]
- A distal dimerization domain is essential for DNA-binding by the atypical HNF1 homeodomain. Nucleic Acids Res. 1990;18:5853-63.
- [Google Scholar]
- A novel syndrome of diabetes mellitus, renal dysfunction and genital malformation associated with a partial deletion of the pseudo-POU domain of hepatocyte nuclear factor-1beta. Hum Mol Genet. 1999;8:2001-8.
- [Google Scholar]
- Sucrase-isomaltase gene transcription requires the hepatocyte nuclear factor-1 (HNF-1) regulatory element and is regulated by the ratio of HNF-1 alpha to HNF-1 beta. J Biol Chem. 2001;276:32122-8.
- [Google Scholar]
- Salmon HNF1: CDNA sequence, evolution, tissue specificity and binding to the salmon serum albumin promoter. J Mol Biol. 1995;247:1-10.
- [Google Scholar]
- Expression patterns of vHNF1 and HNF1 homeoproteins in early postimplantation embryos suggest distinct and sequential developmental roles. Development. 1992;116:783-97.
- [Google Scholar]
- The NMR solution structure of the non-classical homeodomain from the rat liver LFB1/HNF1 transcription factor. J Mol Biol. 1997;267:673-83.
- [Google Scholar]
- Genetic analysis of late-onset type 2 diabetes in a mouse model of human complex trait. Diabetes. 1999;48:1168-74.
- [Google Scholar]
- A POU-A related region dictates DNA binding specificity of LFB1/HNF1 by orienting the two XL-homeodomains in the dimer. EMBO J. 1992;11:4119-29.
- [Google Scholar]
- vHNF1 is expressed in epithelial cells of distinct embryonic origin during development and precedes HNF1 expression. Mech Dev. 1991;36:47-58.
- [Google Scholar]
- Variant hepatocyte nuclear factor 1 is required for visceral endoderm specification. Development. 1999;126:4795-805.
- [Google Scholar]
- Expression profile of MODY3/HNF-1alpha protein in the developing mouse pancreas. Diabetologia. 2002;45:1142-53.
- [Google Scholar]
- HNF-1 alpha and HNF-1 beta (vHNF-1) share dimerization and homeo domains, but not activation domains, and form heterodimers in vitro. Genes Dev. 1991;5:1042-56.
- [Google Scholar]
- HNF1beta/TCF2 mutations impair transactivation potential through altered co-regulator recruitment. Hum Mol Genet. 2004;13:3139-49.
- [Google Scholar]
- Expression of the vHNF1/HNF1beta homeoprotein gene during mouse organogenesis. Mech Dev. 1999;89:211-3.
- [Google Scholar]
- Lack of TCF2/vHNF1 in mice leads to pancreas agenesis. Proc Natl Acad Sci U S A. 2005;102:1490-5.
- [Google Scholar]
- Developmental regulation and tissue distribution of the liver transcription factor LFB1 (HNF1) in Xenopus laevis. Mol Cell Biol. 1993;13:421-31.
- [Google Scholar]
- Cloning and developmental expression of LFB3/HNF1 beta transcription factor in Xenopus laevis. Mech Dev. 1994;47:19-28.
- [Google Scholar]
- The mutated human gene encoding hepatocyte nuclear factor 1beta inhibits kidney formation in developing Xenopus embryos. Proc Natl Acad Sci U S A. 2000;97:4695-700.
- [Google Scholar]
- A molecular link between the common phenotypes of type 1 glycogen storage disease and HNF1alpha-null mice. J Biol Chem. 2001;276:7963-7.
- [Google Scholar]
- Molecular mechanisms and clinical pathophysiology of maturity-onset diabetes of the young. N Engl J Med. 2001;345:971-80.
- [Google Scholar]
- Genetic basis of maturity-onset diabetes of the young. Endocrinol Metab Clin North Am. 2006;35:371-84, x.
- [Google Scholar]
- Hepatocyte nuclear factor 1 inactivation results in hepatic dysfunction, phenylketonuria, and renal Fanconi syndrome. Cell. 1996;84:575-85.
- [Google Scholar]
- Roles of HNF-1beta in kidney development and congenital cystic diseases. Kidney Int. 2005;68:1944-7.
- [Google Scholar]
- Glomerulocystic kidney disease in mice with a targeted inactivation of Wwtr1. Proc Natl Acad Sci U S A. 2007;104:1631-6.
- [Google Scholar]
- Regulation of kidney-specific Ksp-cadherin gene promoter by hepatocyte nuclear factor-1beta. Am J Physiol Renal Physiol. 2002;283:F839-51.
- [Google Scholar]
- Pathophysiological regulation of renal SLC22A organic ion transporters in acute kidney injury: Pharmacological and toxicological implications. Pharmacol Ther. 2010;125:79-91.
- [Google Scholar]
- Transcriptional regulation of human and mouse organic anion transporter 1 by hepatocyte nuclear factor 1 alpha/beta. J Pharmacol Exp Ther. 2008;324:784-90.
- [Google Scholar]
- Regulation of tissue-specific expression of renal organic anion transporters by hepatocyte nuclear factor 1 α/β and DNA methylation. J Pharmacol Exp Ther. 2012;340:648-55.
- [Google Scholar]
- Essential role for collectrin in renal amino acid transport. Nature. 2006;444:1088-91.
- [Google Scholar]
- Aminoaciduria and altered renal expression of luminal amino acid transporters in mice lacking novel gene collectrin. Am J Physiol Renal Physiol. 2007;292:F533-44.
- [Google Scholar]
- The role for HNF-1beta-targeted collectrin in maintenance of primary cilia and cell polarity in collecting duct cells. PLoS One. 2007;2:e414.
- [Google Scholar]
- Mutation in hepatocyte nuclear factor-1 beta gene (TCF2) associated with MODY. Nat Genet. 1997;17:384-5.
- [Google Scholar]
- Frameshift mutation, A263fsinsGG, in the hepatocyte nuclear factor-1beta gene associated with diabetes and renal dysfunction. Diabetes. 1998;47:1354-5.
- [Google Scholar]
- Abnormal nephron development associated with a frameshift mutation in the transcription factor hepatocyte nuclear factor-1 beta. Kidney Int. 2000;57:898-907.
- [Google Scholar]
- Mutations in hepatocyte nuclear factor-1beta and their related phenotypes. J Med Genet. 2006;43:84-90.
- [Google Scholar]
- Hepatocyte nuclear factor-1beta: A new kindred with renal cysts and diabetes and gene expression in normal human development. J Am Soc Nephrol. 2001;12:2175-80.
- [Google Scholar]
- Identification of a gain-of-function mutation in the HNF-1beta gene in a Japanese family with MODY. Diabetologia. 2002;45:154-5.
- [Google Scholar]
- HNF1B mutation in a Turkish child with renal and exocrine pancreas insufficiency, diabetes and liver disease. Pediatr Diabetes. 2012;13:e1-5.
- [Google Scholar]
- A novel hepatocyte nuclear factor-1β (MODY 5) gene mutation in a Romanian boy with pancreatic calcifications, renal and hepatic dysfunction. Georgian Med News. 2011;193:55-60.
- [Google Scholar]
- The renal cysts and diabetes (RCAD) syndrome in a child with deletion of the hepatocyte nuclear factor-1β gene. Indian J Pediatr. 2010;77:1429-31.
- [Google Scholar]
- Phenotype and outcome in hereditary tubulointerstitial nephritis secondary to UMOD mutations. Clin J Am Soc Nephrol. 2011;6:2429-38.
- [Google Scholar]
- A 17q12 chromosomal duplication associated with renal disease and esophageal atresia. Eur J Med Genet. 2011;54:e437-40.
- [Google Scholar]
- Chronic diabetic complications in patients with MODY3 diabetes. Diabetologia. 1998;41:467-73.
- [Google Scholar]
- Recurrent reciprocal genomic rearrangements of 17q12 are associated with renal disease, diabetes, and epilepsy. Am J Hum Genet. 2007;81:1057-69.
- [Google Scholar]
- Recurrent microdeletion at 17q12 as a cause of Mayer-Rokitansky-Kuster-Hauser (MRKH) syndrome: Two case reports. Orphanet J Rare Dis. 2009;4:25.
- [Google Scholar]
- Autism in three patients with cystic or hyperechogenic kidneys and chromosome 17q12 deletion. Nephrol Dial Transplant. 2010;25:3430-3.
- [Google Scholar]
- Mutations in the hepatocyte nuclear factor-1β (HNF1B) gene are common with combined uterine and renal malformations but are not found with isolated uterine malformations. Am J Obstet Gynecol. 2010;203:364.e1-5.
- [Google Scholar]
- Novel molecular pathways in renal Mg2+ transport: A guided tour along the nephron. Curr Opin Nephrol Hypertens. 2010;19:456-62.
- [Google Scholar]
- HNF1B alterations associated with congenital anomalies of the kidney and urinary tract. Pediatr Nephrol. 2010;25:1073-9.
- [Google Scholar]
- HNF-1B specifically regulates the transcription of the γa-subunit of the Na+/K+-ATPase. Biochem Biophys Res Commun. 2011;404:284-90.
- [Google Scholar]




